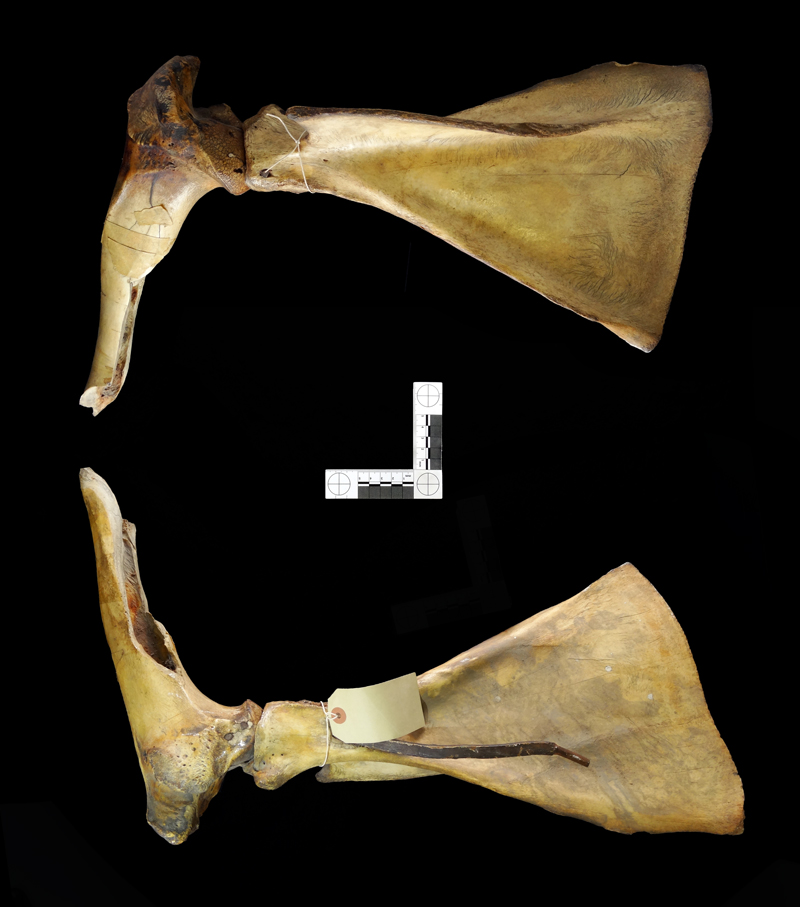
Last Friday I gave you this rather large scapula that I discovered in a crate in the Horniman’s stores to identify:
It wasn’t an easy one, since there are relatively few distinctive features on a scapula compared to something like a skull.
Jake has talked about scapulae on his blog before and that provides a good place to see that this specimen is most likely from an ungulate – but an ungulate much bigger than a Red Deer. This led to suggestions for Cow, Horse, Aurochs and one of the larger species of deer.
Outside the comments section on Zygoma there were also suggestions of Giraffe and Giant Irish Deer and I wondered about Camel.
All in all, there were a lot of suggestions, but none of these looked quite right when I searched for comparative material – although finding good images of scapulae online wasn’t easy. I did, however, find a useful video explaining the differences between Horse, Cow (or Ox) and Camel scapulae:
This was enough for me to rule out each of those animals, although the closest was the Cow – in particular the relative sizes of the two faces (called fossae) on either side of the raised ridge called the spine. However, the shape of the acromion (the hooked bit of the raised spine that points towards the shoulder joint) didn’t seem blunt enough for a Cow.
The size differences in the fossae turn out to be about the same in Sheep and deer as in Cow, which led me back in the direction of Jake’s deer scapulae, which seemed to most closely match the shape, if not the absolute size.
Taking the size into account I realised that this animal must stand almost twice the height of a Red Deer, which narrows it down to just one modern species – the Moose or Eurasian Elk Alces alces (Linnaeus, 1758), which can stand at over 2m at the shoulder compared to the Scottish Red Deer’s (still imposing) 1.22m.
I still need to double-check my identification against a confirmed Moose scapula, but from looking at some images of Moose skeletons online it seems that the shape of both the fossae and the acromion fit well.
So a big thanks to everyone for their help in identifying this and special props to newcomer Jeanie who seems to have been spot-on about this being from a cervid. Thanks!
Last Friday I gave you this big chunk of bone to identify:
I was hoping that it might be a little bit of a challenge because it doesn’t seem to have any really diagnostic characters, but your shape-matching skills were good and several of you managed to get a close identification.
Heather was straight in with the suggestion of it being the back part of the frontal bone (the bit that makes up the front and top of the skull) of an Aurochs – the very large, extinct ancestor of modern Cows. Wouter van Gestel also suggested one of the large bovids – the Asian Water Buffalo, and Ben Gruwier agreed with both Heather and Wouter in saying that it was from a large bovid.
This was as far as I had managed to get with the identification myself, however the specimen had a number (39.16), which I was able to check against the natural history registers. The first part of the number told me to check in the register for the year 1939 and it was the 16th entry for that year, so it was easy to find (unlike with some numbering systems with museum specimens).
It turns out that this specimen is in fact the frontal bone of a Gaur or Indian Bison Bos gaurus Smith,1827, and it turns out that the Gaur has a distinctive ridge between the horns, which is what this specimen is showing, so I should have been able to work it out from the morphology (I will be able to in future).
Bull Gaur can weigh up to 1.5 tonnes and stand 2.2m (7’2″) to the shoulder – they’re enormous. Their only natural predators are the Tigers and large Crocodiles they share their Southeast Asian forest habitat with, but even then Gaur have been known to kill Tigers by trampling and goring them.
Perhaps unsurprisingly these animals are far more risk from humans and have been hunted for meat and trophies until they have become threatened. They are protected by CITES (the Convention on International Trade in Endangered Species), but illegal trade continues and their forest habitats are constantly being lost due to human encroachment.
It’s disheartening that so many of my mystery objects end with a comment about human activities driving a species towards extinction, but unfortunately it’s a massive problem in the world we live in. I wonder if there will be any wild Gaur left in 2039, just 100 years after this specimen was collected?
This Friday I have a big chunk of bone for you to have a go at identifying. It’s something I came across in a stray box – I expect it belongs to another specimen, but I need to identify what species it might belong to before I can start checking. Any ideas?
Your suggestions will be most welcome in the comments section below – let’s see if we can work this out.
Last Friday I gave you this object to have a go at identifying:
It was not an easy one and I was hoping that nobody would work out what it was, but as usual some of you managed to figure it out. So, big congratulations to cackhandedkate and Jake who suggested the surface of a tongue and henstridgesj for working out that these 2mm hooks are from the tongue of a cat – a BIG cat.
These are in fact the barbs or aculei from the tongue of a young Tiger Panthera tigris (Linnaeus, 1758)
This small sealed box has no date associated, but the style of the label and the number suggest that it probably one of the early specimens acquired by Frederick Horniman before he built the Horniman Museum, dating to 1886 or perhaps earlier.
The aculei on cat tongues are interesting adaptations not seen in other carnivores (at least none that I can find any information for). The rows of hooks are ideal for grooming – like a stiff brush, but they are also a useful tool for removing meat from bones.
This is particularly handy when you have a relatively short face with incisors that form a broad straight row for clamping windpipes shut, unlike the narrower incisor row that you find in the dogs, which act a bit like like tweezers for removing meat on bone.
The aculei are made of keratin, the same protein that claws, hair and horns are made from. It may seem quite difficult to evolve lots of small claws on your tongue, but you might be surprised to know that the cells that secrete keratin (called keratinocytes) are the the most common cells on the surface of your skin (including your tongue) where they play an important role in fighting infection and repairing damage.
In order for these cells to secrete enough keratin to grow a small claw, they need to get a bit bigger so they can secrete more of the protein. They also need a simple mechanism that gives the resulting structure a useful shape.
Mammalian tongues are already covered in little structures called ‘papilla’, with three types containing taste buds and one type, the filiform papilla, that provide grip on the surface of the tongue, making it easier to eat ice-cream. It’s these filiform papilla that have adapted in the cats to make a structure with enough grip to lick the meat off a bone.
I’m actually a bit surprised that more mammals don’t have tongue barbs like cats, although there are other animals out there that have specialised tongues with other keratinous structures, like the horny tips of Woodpecker tongues and the tiny bristles of some Fruitbats.
Still, nothing says ‘obligate carnivore’ like a tongue covered in sharp hooks. Considering the length of the aculei I wouldn’t fancy being licked by a Tiger – their tongue looks like it could take human skin right off. However, it has given me an idea for how to quickly and easily remove wallpaper using a Tiger and a bucket or two of Bovril.
This week I have a mystery object that’s a bit dusty and not much to look at, but which is one of my favourite historic specimens at the Horniman Museum.
I have the feeling that it might stump everyone this time, but let’s see how you do. Any idea what this is?
Scale = 10mm
As usual you can put your comments, questions and suggestions below and I’ll give you some clues if you need them. Good luck!
Next week we’ll be reviewing our mineral collections at the Horniman, which means that I’ve been trying to track down the various minerals in the collection. Amongst them I found this rather lovely sample of bismuth – a non-toxic, silvery, heavy metal that can naturally grow into an intricate step-structure crystal.
As the outer surface of the bismuth crystal oxidises it becomes iridescent, creating a beautiful and multifaceted structure that is really quite gorgeous. Hope you like it!
It was a bit of a mean one, since it had no scale bar and the specimen is quite old and dried out, so it doesn’t look much like the living animal.
I had hoped that this would mean that nobody would manage to identify it, but I wasn’t at all surprised when correct suggestions started coming in.
Dave Hone was the first to get the correct kind of animal, although he was a bit thrown by the outer surface – vannabarber was also on the right track, but thrown by the texture. In fact the texture led to some interesting suggestions, including pumice, fossil, bezoar and Pompeian pinecone.
In the end, henstridgesj made the right connection and identified the species, with Anna Pike, rachel and Crispin Wiles all coming to the same conclusion. This is the dried and shrivelled remnant of a Gumboot (or Giant Western Fiery) Chiton Cryptochiton stelleri (Middendorff, 1847). Also known as the ‘Wandering Meatloaf’ for obvious reasons!
Chitons are an ancient class of mollusc called the Polyplacophora – a name that means “bearing many (or several) tablets (or plates)”. They get this name from the eight plates (also known as valves) that they have on their backs.
Most chitons have these valves visible (see below), but the huge Gumboot Chiton has the valves hidden underneath their rubbery girdle.
Tonicella lineata showing the eight valves characteristic of chitons
Chitons are remarkably conservative animals, having changed little since the group arose around half a billion years ago. They have few predators and manage to live a blameless and slow-paced life feeding on algae and detritus on rocks in the world’s oceans, that they rasp off with a fairly simple rasping radula.
There are few ways of spending time on the sea shore that are more enjoyable than turning over rocks in the quest for chitons. Except maybe finding washed-up bones. Or maybe finding both together!
However, once again I feel as though I’m in a similar situation, especially with regard to the time I have available for writing answers. Therefore, I have decided to have a go at making the mystery object fortnightly, the answers being published a full week after the mystery object.
This longer turn-around for the answer will let me open the mystery object up for more guest contributions (since getting the answer over a weekend is usually a struggle) and hopefully that will make it more fun for everyone!
On to the object:
Do you have any idea what this might be?
Leave your questions, comments and suggestions below – preferably in a cryptic format so others can have a go.
And don’t forget, the answer will be posted next Friday. Enjoy!
On Friday I gave you this rather beautiful object to identify,which came to light during our mollusc Bioblitz last week:
It turns out that it didn’t prove much of a challenge and was identified to species level in no time. So well done to Kevin, Anna Pike, @benharvey1 and Carlos Grau!
In fact, Carlos went a step further than identifying the specimen and told the very story I was planning to tell in this post. It’s great to hear stories like this about specimens or species, so I’ll share it with you in Carlos’ words:
This picture immediately brought back memories of my old seashell-collecting guide I had when I was about 12 and haven’t looked at for years and years (I will look for it next time I’m at my parent’s). The book said that this species was considered so valuable that fakes were made in rice paste by Chinese artisans, and that the counterfeits are now more rare and valuable than the actual shell! I remember finding that bit of information amazing.
It’s been so long I had to Google the book, it’s “Guide to Seashells of the World” by R. Tucker Abbott.
This post will only be short, because other people have been dealing with this issue perfectly well without the need for my input, but I thought it might be useful to make visible the outcome of an extended conversation I had with AAH proponent Marcel F. Williams, in the comments section of my earlier post.
I decided to check some independent evidence about an inference made by Alister Hardy that led to the development of the AAH in the first place – the idea that having layers of subcutaneous fat was a trait unique to humans and aquatic mammals. This idea is still regularly cited by AAH proponents (especially Elaine Morgan) as a line of evidence for evolutionary convergence between humans and marine mammals due to the sharing of an aquatic habitat.
However, on checking the literature on primate husbandry it turns out that other primates have levels of subcutaneous fat that are directly comparable to humans if the animals have a ready supply of food, suggesting that humans are no different to other primates except in having a more stable food supply and leading a more sedentary life. This is supported by data from modern hunter-gatherer groups, who exhibit far lower fat levels than either farming or Western populations. Here’s a summary of the data with a link to the research:
In short, humans are by no means unique in the primates with regard to their proportion of subcutaneous fat, so if any AAH proponent pulls out that old chestnut in conversation, be sure to put them right.
On Friday I gave you these two objects (with a third photo to show the end) to identify.
The specimens had me a bit stumped. They are keratinous (keratin is the protein that makes up fingernail, hair and horn amongst other things) and perhaps unsurprisingly they had been labelled as “Artiodactyla horns” given their overall shape. Of course, if they were artiodactyl horns they would be from a bovid (antelope, sheep, cow, etc.), since the other artiodactyls don’t have unbranching horns.
There are nearly 150 bovid species, ranging from the gigantic Gaur to the miniature Royal Antelope, and so far I’ve not been able to find any with horns quite like this. Moreover, the very small size, pattern of growth and relatively shallow depth of the inside chamber of the sheath, don’t really agree with the identification.
My next thought was the spur of a galliform bird, like a Chicken. I compared these specimens to the spur of a male Chicken skeleton in the Horniman’s collection and they looked quite different and much too long, so I gave up on that idea.
Finally I started to consider claws of all sorts of animals, but this didn’t make much sense to me as claws aren’t usually round in cross-section and they have wear facets from being used to walk, climb or dig. You don’t have claws this big if you’re not going to use them and I couldn’t think of much use for a claw like this.
So that left me stumped.
Fortunately, Mieke Roth came to my rescue and made me reassess the Chicken spur identification. It turns out that the Chicken I compared the spur to much have had his spur sheaths removed and they’d hardly grown back.
As you can see, the spur identification fits perfectly!
Many thanks to everyone for their suggestions on this mystery object. It really helps to have fresh eyes looking at a problem and suggesting something you’ve discarded in error!
I’ve been running this blog for over four years now and it’s been some time since I really engaged with it beyond posting my mystery object on Friday and the answer on Monday.
I finally forced myself out of bed extra early on a Saturday morning to give the old blog a bit of a spring clean. What I found was both distressing and heartening.
The links I had to other sites were so out of date that half of them had moved or had simply stopped being updated over a year ago.
With others I found that the content had moved on, from blogs about being a hopeful museum newbie to blogs about being a capable museum professional (see Wunderkammer and Museological Meanderings as examples). In the case of Jake’s bones the blog had changed from being a 9-year-old talking about his bone collection, to an eleven-year-old talking about his forthcoming book on bones.
This sort of progression is wonderful to see and it led me to consider how my blog has changed over the years.
An opportunity for self reflection
The photos now are generally of better quality than they were at the outset, so that’s good. However, I feel that the detail and quality of the answers to the mystery objects have deteriorated, as the time I have available to research and write about them has decreased.
The mystery objects themselves have also become a bit less inspiring, perhaps because the number of striking specimens in the Horniman’s collections that need identification has decreased.
One thing is certain however, after nearly 4 years of running the mystery object I have built up a fantastic community of people who come to Zygoma to have a go at identifying the Friday mystery object, which is something worthwhile.
So now I am left with a quandary – how do I rethink Zygoma to make better use of my increasingly limited time, while maintaining the sense of community on the site?
I may try reducing the number of mystery objects so they’re not a weekly feature, but if I do that I can fill the gap with photos of interesting specimens I’ve come across at work and perhaps specimens that people request.
If you have any ideas about what you’d like to see on Zygoma please leave a comment below – I’d also appreciate links to other related blogs that I may have missed when tidying up my list, so please feel free to suggest away!
As I suspected, it wasn’t too much of a challenge. Jake got in quick with the suggestion of a Sri Lankan Python, with Gina Allnatt, Kevin and Rhina Duque-Thues all agreeing with a generic Python sp. identification. Nicola Newton also suggested Python, but she was of the opinion that it was probably a Royal Python or Carpet Python on the basis of the size.
I agree with Nicola and think that this is most likely the skull of a Royal (or Ball) Python Python regius (Shaw, 1802), based on the size of the specimen and the shape of the supratemporal and frontal bones compared to other Pythons I was able to find images for (if you don’t know which bones I mean you can see what I mean on a handy annotated picture of a Python skull from the Digimorph website).
The skull is one of several similar specimens from the old King’s College teaching collection, so it’s a pretty good bet that the skulls came from either research animals or pets. The Royal Python is a fairly small African Python that is commonly sold as a pet because it has a mild temperament – so that helps offer a bit of support for the identification, although what’s really needed is a detailed identification key for Python skulls or access to good comparative material.
The lack of good resources for identifying the skulls of snakes is a bit frustrating. After working with bird and mammal skulls, where there are some amazing resources like Skullsite and the Mammalian Crania Photographic Archive I’m still searching for something similar for reptiles. Perhaps the Deep Scaly Project will deliver the goods one day… here’s hoping!
If you’re not familiar with the AAH it basically suggests that human ancestors passed through a semiaquatic stage which provided the selective pressure that has led to the differences seen between humans and other primates. Some people call it the Aquatic Ape Theory, but it lacks the necessary scientific support to be considered a theory so it remains a hypothesis [see comments for discussion of the terminology].
The idea was first suggested by pathologist Max Westenhöfer in 1942 and the first line of evidence in support of the hypothesis was proposed by marine biologist Alister Hardy in 1960. Hardy noted that subcutaneous fat is unusual in terrestrial mammals and is normally associated with marine mammals – raising the very good question ‘why do humans have subcutaneous fat?’ (the answer being because we eat too much and exercise too little – just like some lab monkeys).
The baton was then picked up by writer Elaine Morgan who has championed the AAH since 1972. Here’s Elaine in action on a TED video from 2009:
Elaine Morgan is a great communicator and she’s done a remarkable job of delivering the AAH to a wide audience, but I have concerns that the packaging is more impressive than the contents, from a scientific perspective.
In the video Elaine does a cracking job of setting up the AAH in opposition to the more established Savanna Hypothesis (SavH), which suggests that humans diverged from other primates as a result of exploiting more arid environments. She then suggests that the SavH has been discounted on the basis of palaeoenvironmental data, leaving a paradigm gap that should (she suggests) be filled by the AAH.
But of course, a paradigm gap should only be filled by a robust theory and when it comes to plotting evolutionary trajectories there is not solid theoretical foundation on how to do it, beyond relying on the physical evidence provided by the fossil record.
In this case that would require fossils of human ancestors to be found in primarily aquatic deposits, something which we do not see, which is surprising, since aquatic environments are usually far better for fossil preservation than terrestrial environments. In fact, taphonomy suggests that early hominid fossils would be more common if the individuals were living and dying in water with any frequency.
Without having physical data in the form of fossils linking hominids to water, it becomes difficult to make a connection without falling back on evolutionary ‘just-so stories‘, that try to explain an observation by relying on a plausible narrative.
The trouble with this is that the public and media love a good narrative, but it simply isn’t scientific unless it can be falsified. I think this is the part of the process that Elaine Morgan doesn’t quite grasp – she is convinced by her own narrative and believes in the hypothesis, but for a scientist it is more appropriate to subscribe to none of the available hypotheses if they cannot provide factual evidence in support. This is where I currently stand.
I am certainly not convinced by statements like:
“Docosahexaenoic acid (DHA) is an omega-3 fatty acid that is found in large amounts in seafood,… It boosts brain growth in mammals. That is why a dolphin has a much bigger brain than a zebra, though they have roughly the same body sizes. The dolphin has a diet rich in DHA. The crucial point is that without a high DHA diet from seafood we could not have developed our big brains. We got smart from eating fish and living in water.” [Quote attributed to Dr Michael Crawford]
This statement starts by comparing two utterly different species, with a very different evolutionary history and current mode of life, then offers a single dietary explanation for the difference in brain size. This is patently nonsense.
If a seafood diet is the main driver of large brain size then the relatively very large brains of Chimpanzees and other Apes become a remarkable oddity and the relatively small brains of Blue Whales become bizarre variants. Moreover, what about people that don’t have access to seafood? Are they unable to grow large brains? No. Clearly there is more going on.
Brain size is not directly linked to a single dietary chemical, it is linked to managing interactions in and with a complex environment – keeping track of seasonal and spatial variation in fruiting or schooling of fish, coordinating group efforts and understanding prey behaviour to hunt more effectively, or problem solving to access food sources that are hard to acquire. Where a big brain provides a selective advantage, it will evolve.
I don’t want this post to turn into a refutation of the AAH, since there’s a lot to say and much has been done elsewhere. What I do want this post to do is highlight that a scientific theory needs to be testable, it needs to consider contradictory information and it needs to be aware of confirmation bias.
The AAH relies strongly on observed similarities in condition between humans and aquatic mammals, but it dismisses other similarities out of hand. For instance, Naked Mole-rats are simply dismissed by Morgan as an example of a non-aquatic mammal that has lost body hair, but they provide evidence that hair loss can occur for reasons beyond aquatic adaptations – which is worthy of note.
It is also worth considering the supporting examples in the context of phylogeny and physiology, which doesn’t seem to happen often. For instance, the Cetacea, walruses and Sirenia are examples of naked aquatic mammals cited in the AAH, but both the Sirenia and walruses retain a short coat of hairs, quite different in structure to the fine body hair of humans. Whales and Sirenia have also been adapting to an aquatic habitat for 50 million years and the ‘nakedness’ of modern examples may be more related to the evolution of large body size and the benefits to thermoregulation provided by mass – which is supported by the fact that the largest species in the Pinnipedia (like the Elephant Seal and Walrus) are much less reliant on fur than the smaller species of seal.
Unless our ancestors were massive, it seems unlikely that they would have been losing their hair in order to survive better in the water.
Of course, that’s not to say that our ancestors avoided water – far from it. Marginal environments are rich sources of food and most terrestrial animals that live near water will exploit it in some way or another. I’m sure our ancestors would have done the same, I’m just unsure about how immersive and influential that exploitation was on our evolutionary trajectory.
So far I am unconvinced by the AAH and the more bad science and overstated arguments I see in support of it, the less convinced I become. Let’s see if any good supporting science with hard facts emerge from the conference in May.
On Friday I asked you to help me identify this mystery object:
Many thanks to everyone who made suggestions about what this sacrum and caudal vertebrae (the bones that make up the tail) could be from. There were some useful ideas that I intend to follow up on, but alas I must apologise to you all because I’ve still not managed to make a confident identification (as yet).
I always struggle with the identification of vertebrae without having good comparative material. Skulls are straightforward to identify as they tend to contain lots of diagnostic features, but with vertebrae there are fewer distinctive feature that allow a straightforward species level identification.
In this case I’m still not sure whether this tail is from a marsupial, a monkey or a mustelid, although I’m pretty certain it’s from a mammal.
I think I may have to mop up some of the loose ends of mystery objects that I’ve not been able to confidently identify at some point, by making a trip to another museum with a bigger collection of postcranial material than I have available at the Horniman.
Of course, sometimes you just have to accept that there isn’t enough information associated with a specimen to make a confident identification at all. Sometimes you also need to ask whether it’s worth the extra time and resources trying to get a good identification for a specimen with no other good data about where, when and by whom it was collected.
This kind of information can turn an interesting display or teaching specimen into an even more useful research specimen, that can be used to address questions about species distribution, population genetics and evolution – amongst other things.
With a specimen that lacks these kinds of data – and which isn’t particularly visually exciting for display – it becomes more difficult to justify going to special efforts to identify it. Nonetheless, I know this specimen will bug me until I work out what it is!





















.svg)


